Dr. Burgert Senekal, University of the Free State
Ensovoort, volume 44 (2023), number 11: 3
Abstract
The study of European populations has a long history, with scholars tracing the movements of peoples and cultures for centuries. However, recent advances in archaeogenetics have breathed new life into this field of research, allowing scientists to investigate the genetic history of ancient populations in unprecedented detail. The study at hand takes a multidisciplinary approach to this topic, drawing on archaeogenetic research, linguistics, and archaeology to trace the distant origins of the Afrikaner.
The study begins by exploring major migrations into Europe, starting from the Palaeolithic era and continuing through to more recent times. By incorporating genetic research from these periods, the current study highlights the various waves of migration that have shaped the genetic makeup of modern European populations. Alongside genetic evidence, the study also considers the physical and cultural remnants of these migrations, including language, diet, stature, symbols, and aspects of belief.
Through this multidisciplinary approach, the current study paints a detailed picture of the development of European and Afrikaner culture over the course of thousands of years. Rather than simply looking at the past few centuries, this study shows that the roots of Afrikaner culture can be traced back much further.
Keywords: Afrikaner; archaeogenetics; culture; Europeans; Indo-Europeans; language; Yamnaya
Introduction
Almost every history of the Afrikaner starts around 1652, when Jan van Riebeeck established a refreshment station at the Cape. However, as Anthony (2007:3) argues, people carry the traces of their origins in their genes, and these have a much larger effect on people than what is commonly recognised,
When you look in the mirror you see not just your face but a museum. Although your face, in one sense, is your own, it is composed of a collage of features you have inherited from your parents, grandparents, great-grandparents, and so on. The lips and eyes that either bother or please you are not yours alone but are also features of your ancestors, long dead perhaps as individuals but still very much alive as fragments in you. Even complex qualities such as your sense of balance, musical abilities, shyness in crowds, or susceptibility to sickness have been lived before. We carry the past around with us all the time, and not just in our bodies. It lives also in our customs, including the way we speak. The past is a set of invisible lenses we wear constantly, and through these we perceive the world and the world perceives us. We stand always on the shoulders of our ancestors, whether or not we look down to acknowledge them.
The current study investigates the distant origins of the Europeans and the Afrikaner in particular. The study integrates recent archaeogenetic research, findings by historical linguistics and archaeology, and focuses on the contemporary effects of Europeans’ distant past. The purpose is to show how the very distant past shapes lived experience in the present, and how culture is not the product of development over decades, or even centuries, but rather millennia.
The genetic origins of the Indo-Europeans
The origins of European inhabitants have been investigated for centuries, but advances in archaeogenetics over the past decade were able to shed new light on the distribution of populations. I focus on archaeogenetic research in the current section.
Anatomically modern humans emerged from Africa less than 60 000 years ago, and settled amongst other places in Europe alongside Neanderthals (Serrano, Ordóñez and Fregel, 2021:180; Posth et al., 2023:117). These early humans intermingled with Neanderthals, and Neanderthal ancestry is shared among all known non-African modern human populations (Skoglund and Mathieson, 2018:384). Admixture between anatomically modern humans and Neanderthals occurred around 53 000 BC (Serrano, Ordóñez and Fregel, 2021:180) and Neanderthal genes contribute between 1 and 3% of the genome of non-African humans (Fu et al., 2016:201; Skoglund and Mathieson, 2018:384; Mallory, Dybo and Balanovsky, 2019:1477; Hanel and Carlberg, 2020:867). The non-African human population further diverged between 53 000 and 43 000 BC into the Western Eurasian Upper Paleolithic lineage and the East Asian/Oceanian lineage (Skoglund and Mathieson, 2018:384; Serrano, Ordóñez and Fregel, 2021:180) (according to Lazaridis (2018:22), this split occurred around 37 000 to 34 000 BC). The East Asian/Oceanian lineage further intermingled with the Denisovans (Skoglund and Mathieson, 2018:384), with Denisovan ancestry contributing between 5 and 8% of the genes of the people of New Guinea (Mallory, Dybo and Balanovsky, 2019:1478).
European genetic history shows a number of major migrations. The first European settlement followed the initial expansion from Africa and by 43 000 to 40 000 BC, anatomically modern humans were already widely distributed in Europe (Skoglund and Mathieson, 2018:388). Fu et al. (2016:203) found that from 35 000 BC onwards, modern Europeans share at least some ancestry with these Paleolithic Europeans.
A major migration into Europe occurred around 33 000 – 28 000 BC and was probably connected to the Aurignacian culture’s replacement with the Gravettian in western Europe (Fu et al., 2016:204; Skoglund and Mathieson, 2018:389; Serrano, Ordóñez and Fregel, 2021:180). However, ancestral lines connected to the Aurignacian did not disappear and resurfaced around 17 000 BC (Fu et al., 2016:204).
Around 12 500 BC, the previous inhabitants of Europe were replaced with a population that would come to form the Mesolithic populations of Europe (Skoglund and Mathieson, 2018:389; Serrano, Ordóñez and Fregel, 2021:180; Posth et al., 2023:124). This was the Western Hunter Gatherer (WHG) population, also known as the Villabruna population (Mittnik et al., 2017:2; Lazaridis, 2018:22), which originated in the Near East (Posth et al., 2023:124). By 11 700 BC, this population would reach as far as Scandinavia (Serrano, Ordóñez and Fregel, 2021:181). Here, the Mesolithic Scandinavian hunter-gatherer gene pool (SHG) was formed by the mingling of the Eastern European Hunter Gatherer (EHG) and WHG tribes from Western Europe (Mittnik et al., 2017:2; Serrano, Ordóñez and Fregel, 2021:181). The EHG population itself was formed in Russia from a mixture of WHG and Upper Paleolithic Siberian ancestry (Lazaridis, 2018:22).
Up until 6 500 BC, Europe was mostly occupied by small settlements of hunter gatherers (Mittnik et al., 2017:2; Racimo et al., 2020:8989). Around 6 500 BC, however, a new population, connected to that found in Neolithic northwest Anatolia and to early farming people of the Levant and northern Iran, entered Europe (Lazaridis, 2018:22; Skoglund and Mathieson, 2018:389; Serrano, Ordóñez and Fregel, 2021:181; Chintalapati, Patterson and Moorjani, 2022:9). This new population spread throughout Europe, with direct evidence of its presence in Iberia in 5 300 BC, in Ireland in 3 100 BC, and in Scandinavia in 2 900 BC (Skoglund and Mathieson, 2018:389) (see also similar dates identified by Allentoft et al. (2022:8) and Chintalapati et al. (2022:14)). In central Europe, the admixture of Neolithic Anatolian farmers and WHG populations resulted in the formation of the Linearbandkeramik (LBK) culture (Serrano, Ordóñez and Fregel, 2021:182). By 2 500 BC, almost all European populations were admixed between these two ancestries (Skoglund and Mathieson, 2018:389). Chintalapati et al. (2022:9) found that the Anatolian farmers contributed between 40% and 98% to the ancestry of Neolithic Europeans.
A further significant change in the European gene pool occurred around the third millennium BC, which was the result of the migration of people from the Yamnaya culture (also spelled Jamnaja) – a subgroup of the Kurgan culture (Mallory, Dybo and Balanovsky, 2019:1474) – from the Pontic-Caspian steppe in southern Russia and Ukraine into Europe (Kristiansen et al., 2017:336; Olalde et al., 2018:1; Skoglund and Mathieson, 2018:389; Racimo et al., 2020:8989; Serrano, Ordóñez and Fregel, 2021:182; Allentoft et al., 2022:8). This steppe population is itself a mixture of populations that were related to Mesolithic Eastern European hunter-gatherers (EHG), Mesolithic Caucasus hunter-gatherers (CHG), as well as to populations from Neolithic and Copper Age Iran (Jones et al., 2015:5; Lazaridis, 2018:24; Skoglund and Mathieson, 2018:389; Kristiansen, 2020:158) (Lazaridis et al. (2022:5) found predominantly Caucasus, Anatolian and Levantine ancestry for the Yamnaya culture, while Jones et al. (2015:5) found predominantly CHG ancestry for the Yamnaya). Chintalapati et al. (2022:15) found that the Yamnaya culture can be distinguished genetically by around 4 400 to 4 000 BC, around a millennium earlier than the dates for this culture given by the archaeological record (3 300 and 2 500 BC (Shishlina, Sevastyanov and Hedges, 2012:179; Iversen and Kroonen, 2017:515; Anthony, 2023:14)). The Yamnaya population also spread eastwards to found the Afanasievo culture in Siberia, and they also reached India (Lazaridis, 2018:24).
Steppe ancestry appeared in southeastern Europe by 4 000 BC, northeastern Europe around 3 000 BC, and central Europe around 2 600 BC (Skoglund and Mathieson, 2018:389). Chintalapati et al. (2022:15) date the migration into Europe to around 3 200 BC. Allentoft et al. (2022:8) also found that beginning 2 200 BC, Yamnaya-related ancestry was already dominant in individuals from Britain, France and the Iberian peninsula. The Yamnaya herders replaced around 75% of central Europeans’ Neolithic ancestry (Haak et al., 2015:5; Serrano, Ordóñez and Fregel, 2021:183) and 90% of the ancestry of people from the British Isles (Chintalapati, Patterson and Moorjani, 2022:13). Present-day Europeans derive between 10% and 60% of their ancestry from Steppe pastoralist-related populations (Chintalapati, Patterson and Moorjani, 2022:12).
Present-day western European populations therefore consist mainly of mixtures of three ancestry components, namely Mesolithic hunter–gatherer, Anatolian Neolithic, and Yamnaya (Haak et al., 2015:4; Jones et al., 2015:2; Skoglund and Mathieson, 2018:390). Anatolian Neolithic ancestry is highest in present-day southern European populations such as Sardinians and lowest in northern European populations, whereas Yamnaya ancestry is highest in north-central Europeans and lowest in the southern European populations (Haak et al., 2015:4; Skoglund and Mathieson, 2018:390) (see also similar distributions identified by Allentoft et al. (2022:18)). Margaryan et al. (2020:391) however found that although Viking Age Norwegians show a greater genetic similarity with the Steppe populations, Viking Age Danes and Swedes are genetically often similar to Anatolian Neolithic farmers.
Despite the large-scale population displacement from the Steppe, not all Neolithic populations were replaced. The Basques showed continuity through time with no entrance of the Steppe component to its gene pool in the Late Neolithic/Bronze Age periods (Serrano, Ordóñez and Fregel, 2021:183). The later Picts (300 – 900 AD), however, have been shown to be a mix between contemporary British populations and not a genetically homogenous group (Morez et al., 2022).
The migration in the third millennium BC resulted in the formation of the Corded Ware Culture (2 900 to 2 200 BC) in north-central and northeastern Europe (Kristiansen et al., 2017:336; Racimo et al., 2020:8989; Serrano, Ordóñez and Fregel, 2021:182; Anthony, 2023:13) (which includes the Battle Axe and Single Grave cultures), as well as the Bell Beaker Culture (2 750 – 1800 BC) in western Europe (Olalde et al., 2018:1; Racimo et al., 2020:8989; Serrano, Ordóñez and Fregel, 2021:182; Anthony, 2023:13). The Corded Ware Culture originated through the interactions between migrants from the Yamnaya culture and the existing Neolithic cultures of Europe (Allentoft et al., 2015:171; Kristiansen et al., 2017:342). People belonging to the Corded Ware Culture are therefore closely related to people from the Yamnaya culture (Allentoft et al., 2015:169; Haak et al., 2015:2; Olander, 2019:27; Racimo et al., 2020:8989; Lazaridis et al., 2022:7), as is the case for people belonging to the Bell Beaker culture (Racimo et al., 2020:8989; Chintalapati, Patterson and Moorjani, 2022:13).
Allentoft et al. (2022:16) also conducted a more fine-grained analysis of the genetic structure in southern Scandinavia after the introduction of Steppe-related ancestry, and found three stages in this area’s genetic structure:
- An early stage between 2 600 BC and 2 300 BC, where Scandinavians cluster with early Corded Ware Culture individuals from Eastern Europe, and who have prominent Steppe-related ancestry;
- An intermediate stage where Scandinavians cluster with central and western Europeans; and
- A final stage from c. 2 000 BC onwards, where a distinct cluster of Scandinavian individuals appear.
Allentoft et al. (2022:16) further found that, for later Iron Age and Viking Age Scandinavians as well as ancient European populations outside of Scandinavia with a confirmed Scandinavian or Germanic link, the last stage serves as the primary source.
During the last centuries BC and the first millennium AD, Germanic tribes (amongst others) moved south and settled in various parts of Europe. These migrations are well known from archeological sources, and are also evident in the genetic composition of populations. Stolarek et al. (2019:8) for instance found that, from the second century BC, migrations of Goths into Europe from Southern Scandinavia can be supported through genetic evidence. Margaryan et al. (2020:391) was also able to identify a genetic link between Longobards and individuals from Kärda in southern Sweden. This migration also included the migration to England and from the later Roman period into the eleventh century, a mass migration of Germanic tribes into England is discernible in the genetic record (Gretzinger et al., 2022:118). To the east, Neparáczki et al. (2018) studied population movements into modern day Hungary in the tenth century. The Viking Age also left genetic traces, with Swedes moving into the Baltic, Norwegians into Iceland, Greenland and the Isle of Man, and Danes moving into England (Margaryan et al., 2020:392).
With the close of the Viking Age in the eleventh century, the age of large migrations was over for the European population for almost half a millennium. Starting around 1 500 AD, however, European expansionism distributed Europeans throughout the globe.
One such migration was Europeans settling in South Africa from 1652 onwards. These European settlers were mostly composed of Dutch employees of the Vereenigde Oost-Indische Compagnie (VOC), along with German mercenaries and from 1688, French Huguenots. This migration is also traceable through the genetic record: Hollfelder et al. (2020) studied the Afrikaner’s genetic composition and found a strong northwestern European genetic component (for example, Dutch, German and French), rather than a southern or eastern European component. On average, 95.3% of the Afrikaner’s ancestors are of European origin, with the remaining 4.7% people who were brought to the Cape as slaves in colonial times (3.4%) and from local Khoe-San people (1.3%). Admixture with black Africans is very limited (on average 0.8%), but even then the admixture is traced back to slaves imported from West Africa, rather than admixture with local black Africans. Hollfelder et al. (2020) also found that the Afrikaner groups separately from the South African English population, which shows very little admixture between these population groups. Importantly, however, the admixture between predominantly Dutch, German and French populations occurred in the Cape Colony, and as the admixture of Neolithic Anatolian descendents and Yamnaya settlers resulted in the formation of new cultures, so too did the admixture of these European populations (and to a lesser extent, non-European populations) in the Cape Colony lead to a new culture. In other words, while the Afrikaner is genetically predominantly European, geographically speaking this population developed in Africa.
The above admixture of populations is represented in Figure 1. Colours indicate different periods in a simplified way, with pink referring to the Palaeolithic, green the Mesolithic, olive the Neolithic, purple the Bronze Age, blue the Copper Age, turquoise the Iron Age, and orange the early modern age. Note that this figure visualises the genetic connections discussed in the current article and that populations beyond the current focus are not included.
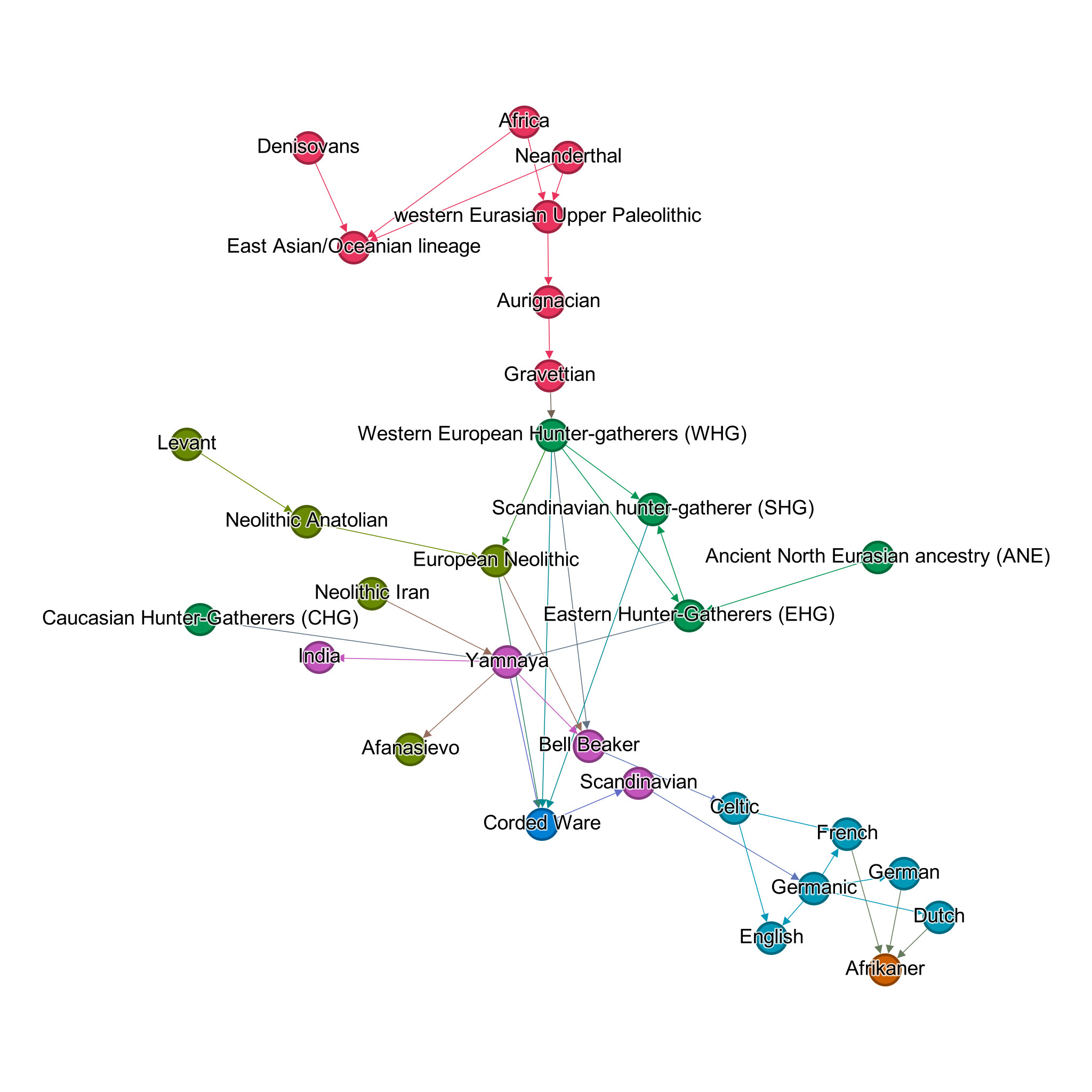
Because Afrikaners have been shown to come mostly from North-Western Europe, and North-Western Europeans can trace their ancestry mostly to the Yamnaya, Neolithic Anatolian and WHG (in this order), it can be expected that the Afrikaner will also trace its lineage back to these three ancestral lines. Unfortunately, no studies have yet been conducted on the Afrikaner’s lineage in terms of Bronze Age and earlier influences.
Remnants of early European migrations in contemporary Europeans, with specific reference to the Afrikaner
Physical characteristics
Many physical traits are heritable, and as such, even population migrations as far back as the Mesolithic have an effect on contemporary populations. Marnetto et al. (2022) studied the effects of WHG, Yamnaya, Neolithic Anatolian and Siberian ancestry on modern populations, and found the following:
-
WHG ancestry is linked to lower cholesterol levels, higher Body Mass Index (BMI), brown hair and light eye colour;
-
Yamnaya ancestry is linked to a strong build, black hair and high-cholesterol;
-
Neolithic Anatolian ancestry is linked to lower BMI, light (but not green) eyes, fair hair, increased age at menarche, and reduced heart rate;
-
Siberian ancestry is linked to dark hair, higher heart rate, lower caffeine consumption, green eye colour and lower age at menarche.
In addition, physical traits such as stature and skin colour are heritable. I focus on these two traits in the subsections below.
Stature
Prior to the industrial revolution, the tallest people in Europe were hunter-gatherers from the Gravettian culture that emerged at least 36 000 years ago, whose males were around 182.7 cm tall on average (in the Mediterranean) (Grasgruber et al., 2014:81). The earliest anatomically modern humans in Europe, around 47 000 to 44 000 BC, had an average male height of 174 cm (Cox et al., 2019:21484). A sedentary lifestyle is associated with a decline in stature, and by the late Neolithic, some European populations were as short as 162 cm (males in the Carpathian Basin). During the third millennium BC, the stature of Europeans again increased to around 169 cm (Grasgruber et al., 2014:82; Cox et al., 2019:21484).
While there are many factors that influence stature, including living standards, health expenditure, and others, diet plays an important role in a population’s average height (Grasgruber et al., 2014). In Grasgruber et al.’s (2014) study, the consumption of meat, dairy, fish and wheat best predicted stature. Grasgruber et al. (2014) also found that lactase persistence (see below) is correlated with stature, although the evidence for this conclusion is weaker than for diet.
In addition, genetics play an important role in stature, with certain genetic lineages more prone to higher stature (Grasgruber et al., 2014; Cox et al., 2019). Neolithic Anatolian farmers (who settled mostly in Southern Europe) were shorter, compared with the taller Yamnaya settlers (who settled mostly in northern Europe), which explains why northern Europeans are today taller on average than southern Europeans (Cox et al., 2019:21489; Hanel and Carlberg, 2020:867). Marnetto et al. (2022:1417) and Skoglund and Mathieson (2018:396) also show that Yamnaya ancestry is linked to height. In addition, Rosenstock et al. (2019) and Cox et al. (2019) found a noticeable increase in stature in the European population around 3 000 BC, which they suggest might be the result of the Yamnaya expansion. Rosenstock et al. (2019:5685) also note a continuation to the present day,
… the stature increase in Later Prehistory was marked enough to establish the Northwest-Southeast gradient in European and Near Eastern stature that can be traced through Antiquity and the Medieval and Modern Period until today.
Today, the tallest population in the world is the Dutch, with an average male height of 183.8 cm (Grasgruber et al., 2014:85).
Myburgh et al. (2017) compared the statures of Dutch, Swiss and South African white males (note: not necessarily Afrikaners) and found average heights of 180.2 cm, 176.5 cm and 178.6 cm, respectively, for the period 1946-1995. For the latest period in their study (1991-1995), Myburgh et al. (2017) do not include average heights for Dutch males, but average heights for Swiss and South African white males are 178.2 cm and 180.5 cm, respectively (the latter is however based on a very small sample). Ngcobo et al. (2021) found a significantly lower average height of white South African males at 175.3 cm, compared to the average height of a black South African male at 170.7 cm. Similarly, Mpeta, Fourie and Inwood (2018) found average heights of white South African males of 174.4 cm for the entire 20th century, compared with 167.6 cm for black males. However, for white South African males born after 1958, the average height is 178.2 cm, compared with 168.9 cm for black males (Mpeta, Fourie and Inwood, 2018). Although no Afrikaner-specific data could be found, it is clear that South African whites are on average taller than other South Africans, which will be influenced by the large Yamnaya genetic component in both the Afrikaner and South African English populations.
Skin colour
Contrary to popular belief, European light skin is not a function of evolution in order to adapt to colder climates. Rather, light skin is the result of genetic factors. The specific combination of pigmentation alleles that is common in western Europe today (SLC24A5 and SLC45A2), and is responsible for lighter skin tones, reached its current high frequency only from 3 000 BC onwards (Skoglund and Mathieson, 2018:394; Hanel and Carlberg, 2020:867). Hanel and Carlberg (2020:867) write that the CHG, EHG and SHG lineages all contain SLC24A5, which suggests that this trait originated around 26 000 – 20 000 BC. As noted earlier, the Yamnaya was composed of CHG and EHG lineages, and since both of these lineages had light skins, the Yamnaya did as well (Hanel and Carlberg, 2020:867). European light skin is therefore partly attributed to the Yamnaya expansion, with Neolithic Anatolian farmers also contributing this trait during their earlier migration, and in addition, the SHG lineage also contributed to light skin (Skoglund and Mathieson, 2018:394; Hanel and Carlberg, 2020:867; Serrano, Ordóñez and Fregel, 2021:181).
Like the South African English population, the Afrikaner is generally considered to be white (Senekal, 2019). This is arguably the Afrikaner’s most controversial aspect, since whiteness was used in the 20th century as the underlying principle behind apartheid. It should be noted, however, that contrary to beliefs that racism developed during the Age of Enlightenment and colonial conquests, as argued by Cox (1959) and repeated by e.g. Maylam (2020), Cole (2015) discusses racial attitudes found among the Norse in the centuries following the Viking Age. The Old Norse term for black Africans was blámenn (singular blámaðr), which literally means “blue men”, and blámenn were never depicted in a positive way in Old Norse literature. Cole (2015:28) writes of the depiction of blámenn in Old Norse literature,
But the blámaðr constitutes the darker side of the Norse conception of Otherness, be it Islamic, Finnic or demonic: a being who does not believe something unnatural, but is something unnatural. He is a creature shaped not by his beliefs, but by the baseness of his blood. (original emphasis)
Cole (2015:31) also links the world of Muspell in the Norse myths to Africa, where Surtr – derived from svartr (“black”) (Cole, 2015:29) – lives. According to Norse mythology, the destruction of the world and the gods will come at the hands of Surtr and the inhabitants of Muspell at the end of the world, Ragnarök.
While whiteness characterises the European population from 3 000 BC onwards, using whiteness as a marker of identity is more difficult to date. However, as Cole (2015) shows, racial thinking was already present in Scandinavian thought by the thirteenth century. It should be noted, however, that the current study considers the origins of Europeans and that the existence of racial thinking among other cultures is not investigated here.
The Indo-European languages
Prior to the publication of recent genomic research, the origins of the Indo-European languages were mainly hypothesised for two regions: the Steppe Hypothesis (Gimbutas, 1956; Mallory, 1989; Anthony, 2007) and the Anatolian Hypothesis (Renfrew, 1989). Iversen and Kroonen (2017:515) and Garnier, Sagart and Sagot (2017:292) claim that the majority of Indo-Europeanists favour the Steppe Hypothesis over the Anatolian Hypothesis, mainly because the combination of historical linguistics and archaeology made this hypothesis the most convincing. According to Olander (2019) and Kroonen et al. (2022), this debate has been decided by genomics research in favour of the former hypothesis. Various authors (Olander, 2019:27; Hanel and Carlberg, 2020:867; Kristiansen, 2020:162; Lazaridis et al., 2022:11) now locate the origin of Indo-European languages with the Yamnaya culture. Lazaridis et al. (2022:11) for instance aver, “All ancient and present-day branches of the Indo-European language family can be derived or at least linked to the early Bronze Age Yamnaya pastoralists of the steppe or genetically similar populations.” However, Kroonen et al. (2022:34) suggest a slight divergence from the Steppe Hypothesis when finding, “the linguistic homeland of the core Indo-European language community cannot be located in the eastern steppe, but must be situated around, and extending to the west of, the Dnieper River.”
Precisely when each branch separated from the Indo-European family tree is still a matter of debate (for a detailed discussion, see Olander (2022)). According to Olander (2019:26), the first branch to separate from the Indo-European language tree is generally considered to be Anatolian, which may have separated between 4 200 and 4 000 BC (because of this early split, Kristiansen (2020) calls Anatolian a sister language of Indo-European). This split was likely followed by the separation of the Tocharian branch, which may have separated around 3 300 BC (Olander, 2019:26). Olander (2019:26) claims that the Italo-Celtic branch is believed to have separated third, possibly between 3 300 and 2 800 BC. The Germanic branch is believed to have separated fourth (Olander, 2019:9), and Proto-Germanic is believed to be linked to the Corded Ware Culture (Olander, 2019:25), although Germanic itself is usually dated to around 500 BC (Bousquette and Salmons, 2017:389; Mallory, Dybo and Balanovsky, 2019:1483).
Kassian et al. (2021) calculated the Indo-European branch separations as in Table 1. Dates BC are given as negatives, and the table is sorted by mean dates. While these dates differ from those offered by Olander (2019:26), Kassian et al. (2021:971-972) claim that these dates largely coincide with established research in linguistics and archaeology.
Table 1. Indo-European branch separations according to Kassian et al. (2021).
|
Event |
Upper |
Lower |
Mean |
|
Anatolian split-off (root) |
-4139 |
-3450 |
-3686 |
|
Tocharian split-off |
-3727 |
-2262 |
-3011 |
|
Inner IE break-up |
-3357 |
-2162 |
-2717 |
|
Balto-Slavic–Indo-Iranian break-up |
-2723 |
-1790 |
-2241 |
|
Italic-Germanic-Celtic break-up |
-2655 |
-1537 |
-2080 |
|
Greek-Armenian break-up |
-2676 |
-1407 |
-2015 |
|
Indo-Iranian break-up |
-2044 |
-1458 |
-1740 |
|
Balto-Slavic break-up |
-1686 |
-855 |
-1250 |
|
Insular Celtic break-up |
-596 |
95 |
-243 |
The Indo-European language family is currently the largest language family in the world, both in terms of the number of speakers and geographical coverage (Kassian et al., 2021:950). Given the size and complexity of the Indo-European language tree, it is not possible to visualise the entire language tree on a single page in such a way that labels remain legible, but Figure 2 shows the position of Afrikaans on this tree. The tree was constructed using the classifications by Ethnologue (https://www.ethnologue.com/), which does not include the extinct Tocharian and Hittite branches.
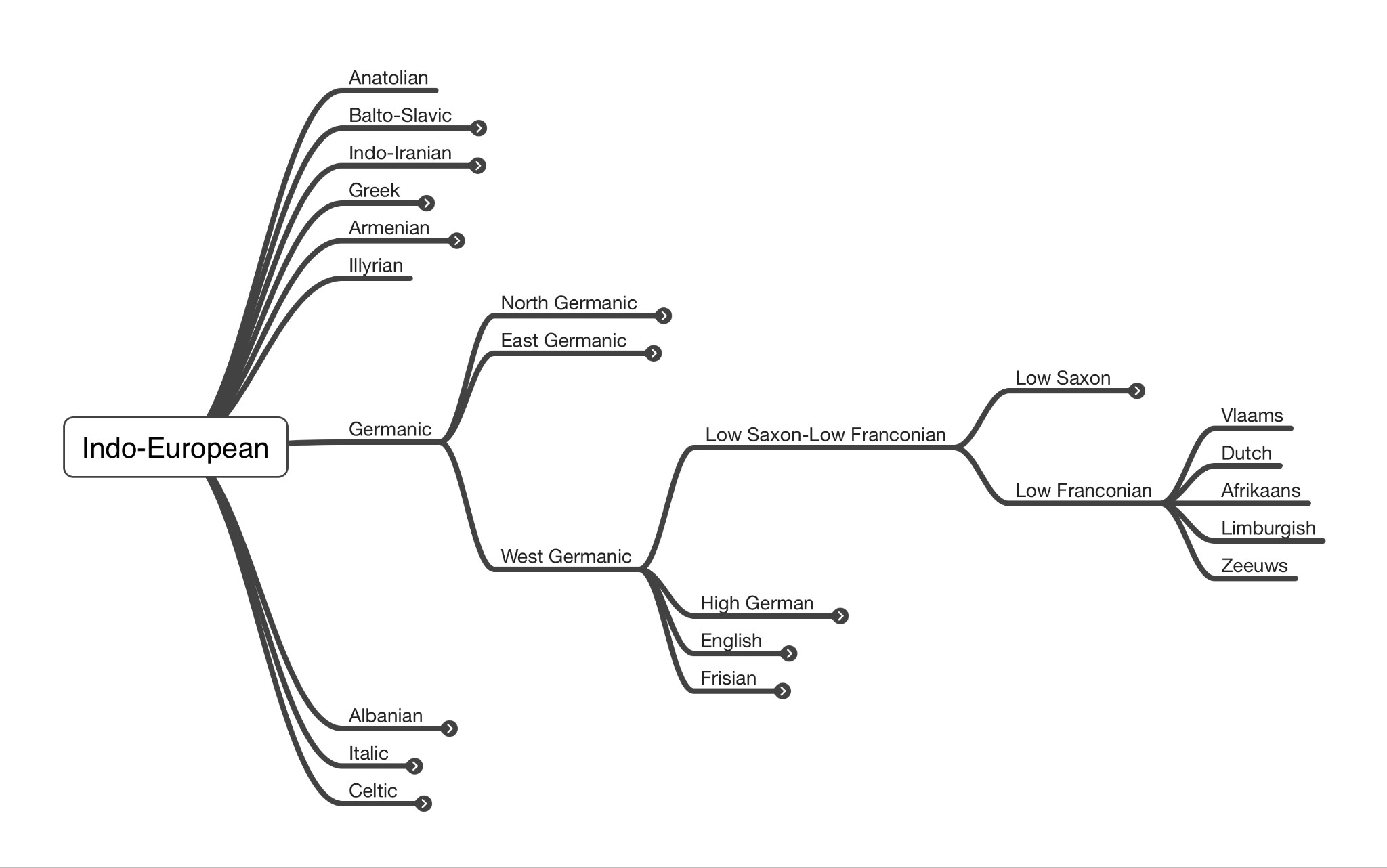
In the most basic definition, Afrikaners are seen as white Afrikaans-speaking people (Senekal, 2019). As discussed above, a light skin colour is inherited to a large extent from the Yamnaya, and the same is the case with language. Afrikaans established itself as the only Indo-European language spoken natively by a culture that originated in sub-Saharan Africa. Semantically and grammatically, Afrikaans is classified as a West Germanic, Low-Franconian language, alongside Dutch, Limburgish, Zeeuws and Flemish. Geographically, however, Afrikaans is an African language, as argued by Willemse (2018) and the South African Constitutional Court (Democratic Alliance, 2021). Afrikaans is therefore – like the Afrikaners themselves – simultaneously European and African, depending on whether lineage or geography is emphasised.
The Yamnaya diet
Iversen and Kroonen (2017) make a case for the hybrid origin of the Germanic languages by postulating that the coexistence of Neolithic and Corded Ware Culture populations between 2 850 BC and 1 500 BC resulted in Neolithic loan words being incorporated into Proto-Germanic. While some Indo-European words exist that relate to crop cultivation, such as for cereal, grain and to plough, important founder crops such as flax, lentils, peas, chickpeas, garlic and turnips have no Indo-European lexical equivalents (Iversen and Kroonen, 2017:516, 518). These and other words were incorporated into Proto-Germanic from Neolithic farming communities and constitute some of the very little evidence for a Neolithic European language (see also Bousquette and Salmons (2017:390)). According to Iversen and Kroonen (2017), the absence of these words indicates that these foods were not known to the carriers of the Indo-European languages, namely the Yamnaya. Rather, Indo-European languages share vocabulary related to dairy production, such as for to milk, buttermilk, sour milk, cow, and cheese (Iversen and Kroonen, 2017:515).
Iversen and Kroonen’s (2017) linguistic research is supported by isotope analysis conducted by Shishlina, Sevastyanov and Hedges (2012). Shishlina, Sevastyanov and Hedges (2012) found that the majority of the Yamnaya diet consisted of animal products (meat and milk), river and lake aquatic products and some wild plants.
Based on the evidence by Shishlina, Sevastyanov and Hedges (2012), Iversen and Kroonen (2017), Garnier, Sagart and Sagot (2017) and Segurel et al. (2020), the Yamnaya diet consisted mainly of dairy, meats and cereal, with some wild plants also consumed. Cheese, which was invented by early farmers in Northwest Anatolia ca. 6 500 BC (Evershed et al., 2008), also formed part of this diet, as was yoghurt (Anthony, 2007:303, 2023:27). Anthony (2023:28) however avers that cereal was not consumed in large quantities. In terms of alcohol, the Yamnaya drank mead (Anthony, 2007:303).
European lactase persistence – the ability to digest lactose and therefore milk in adulthood – has been suggested to be inherited from the Yamnaya culture (Allentoft et al., 2015:171) and is commonly found in European populations from about 1 000 BC (Skoglund and Mathieson, 2018:394). Segurel et al. (2020:6) found that while lactase persistence is found in European genes from 3 950 BC, thus predating the Yamnaya expansion, the expansion of lactase persistence is congruent with the Yamnaya expansion and therefore likely spread by the Yamnaya populations. In another study, Furtwängler et al. (2020:7) found lactase persistence in Switzerland as early as 2 105 – 2 036 BC. Burger et al. (2020:4310), however, found too little evidence of lactase persistence in the European Bronze Age to support the conclusion that this genetic trait was spread by the Yamnaya culture. Nevertheless, milk consumption amongst the Yamnaya has been proven, and it has been suggested that the added protein from dairy products were instrumental in the Yamnaya expansion (Garnier, Sagart and Sagot, 2017; Wilkin et al., 2021). Garnier, Sagart and Sagot (2017:294) for instance argue that lactase persistence gave the early Indo-Europeans a demographic advantage and possibly an increase in physical stature, both of which resulted in a military advantage over earlier farming communities.
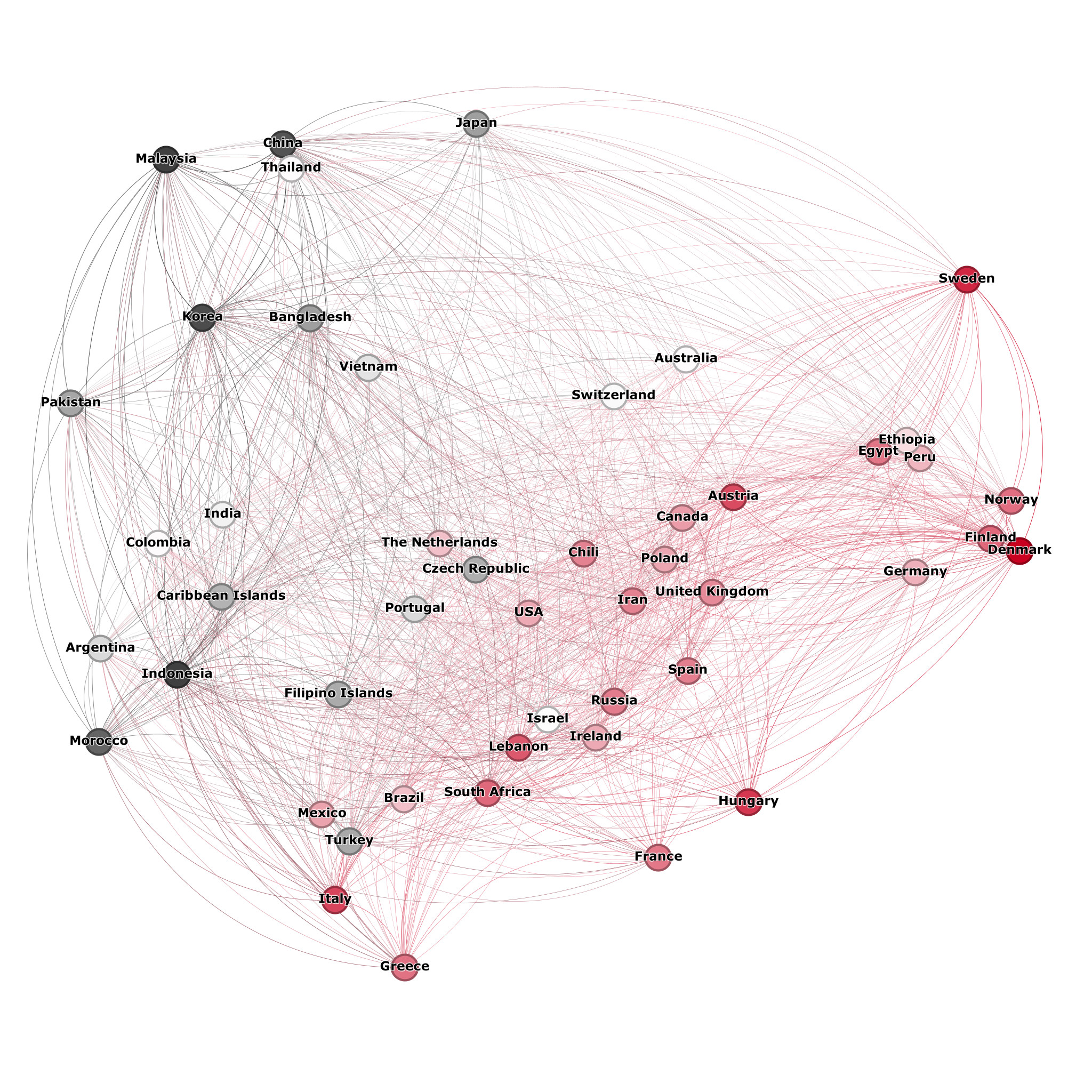
Ancient diets have proved surprisingly persistent in cultures globally. While the spice trade has existed for more than 2 000 years (Pickersgill, 2004:153-155), and global shipping has shipped food across the globe for more than 500 years, studies have shown that regional cuisines remain distinguishable through ingredient combinations (Ahn et al., 2011; Kular, Menezes and Ribeiro, 2011; Zhu et al., 2013; Kim and Chung, 2016; Sajadmanesh et al., 2017; Issa, Alghanim and Obeid, 2018). Of particular note is Ahn et al.’s (2011) finding that a high use of dairy products is characteristic of Western European cooking, since these are also the regions with the highest levels of lactase persistence and Yamnaya ancestry. Senekal (2022) studied Afrikaans recipes, and found that Afrikaans recipes can be considered predominantly Western European, with milk, eggs, cream and butter frequently featured as ingredients. In Senekal (2023), the position of Afrikaans recipes in the global recipe-ingredient network is analysed, where ties are indicated between countries based on the number of ingredients their recipes share. In Figure 3 below (adapted from Senekal (2023)), it is indicated that Afrikaans recipes are similar to European recipes in their relative use of dairy products as ingredients. The graph indicates relative use of dairy products with a scale from black (lowest relative use) to bright red (highest relative use), with white indicating the middle of the spectrum. It can be seen here that most European countries and countries with large European populations use dairy products more frequently than is the case for countries in the far East. This study was based on data gathered from online sources in 2022, which shows the persistence of dairy use in current European recipes.
Cultural and religious vestiges
While the legacy of the WHG, Neolithic Anatolian farmers and the Yamnaya in terms of stature, skin colour, language and diet can be substantiated scientifically, religious and symbolic continuity from the distant past to the present is more difficult to prove, since identifying these ties involves a greater amount of interpretation. I nevertheless discuss a few interesting similarities and hypotheses in the rest of the current article, both as noticed by others and by myself.
As noted earlier, archaeogenetics vindicated the Steppe Hypothesis for the origin of the Indo-European peoples. One of this hypothesis’s greatest proponents, Marija Gimbutas, also suggested that the Indo-European populations brought their own patriarchal beliefs to Europe, which supplanted the existing matriarchal Neolithic beliefs. In her last work, The Living Goddesses (1999), she discusses how vestiges of these Neolithic beliefs were incorporated into Indo-European beliefs, much like Iversen and Kroonen (2017) suggest Neolithic vocabulary was retained in Indo-European languages. Like the Corded Ware Culture was the result of an interplay between Steppe pastoralists and Neolithic farmers (Allentoft et al., 2015:171; Kristiansen et al., 2017:342), so Indo-European beliefs were a combination of the beliefs brought with the Steppe pastoralists and those of the Neolithic farmers.
The Germanic beliefs can serve as an example of this interplay between Neolithic and Bronze Age views. Gimbutas (1999:191) argues that the two distinct families of deities in the Germanic pantheon – the Vanir and the Æsir – represent the coexistence of the Old European and the Indo-European faiths in Germanic tradition. The Æsir (e.g. Óðinn, Þórr, and Týr) are the Indo-European warrior deities of a patriarchal culture, whereas the Vanir (e.g. Freyr, Freyja and Njorðr) are the native Old European deities who are fertile and life-giving. Norse myths note conflict between the two families, but that peace was agreed and ambassadors exchanged, which Gimbutas (1999:191) interprets as an indication of an earlier conflict between the two cultures which has since been resolved.
The Yamnaya was a warrior society, built around “the warrior brotherhood of young men bound by oath to one another and to their ancestors during a ritually mandated raid” (Anthony, 2007:364). These young men were associated with the wolf or dog, with numerous canine teeth worn as pendants found in Yamnaya graves (Anthony, 2007:364). This association with the wolf would later be found amongst Indo-European warrior groups known as the luperci or suodales in Latin, the kouros or ephebes in Greek, fian in Celtic, männerbünde or jungmannschaft in Germanic, and vrātyas or Maruts in Indic (Anthony and Brown, 2017:135). In addition, this association between warriors and dogs resulted in dogs becoming associated with death, and Anthony and Brown (2017:146) note that Greek (Kérberos) and Indic (Śárvara) cognate names for the hellhound were obtained through regular sound changes from a shared Proto-Indo-European root word that meant “spotted.”
Christianity replaced most of the Indo-European beliefs in Europe by around 1 000 AD. Although known as an Abrahamic faith, Christianity however incorporated elements of Indo-European beliefs in its world view. Kehoe (2012) argues that the warrior spirit – inherited in her view from the Yamnaya and developed through e.g. the Germanic tribes – became part of Christianity, which contrasts with the peaceful and non-confrontational teachings of Jesus Christ. Kehoe’s (2012:53) remarks about America would be equally applicable to South Africa,
What would Jesus do if he visited America around 1800? The nation professed Christianity, from time to time in Great Awakening enthusiastic outpourings of piety. But day to day? Slaves were dehumanized, treated like livestock. Social classes were only too obvious in the refinement of the upper and the long hours of labor for the lower. Violence was every day, schoolmasters and parents whipping children, husbands beating wives, drunks brawling, even duels such as the one in which Aaron Burr killed Alexander Hamilton. Indians were being dispossessed still in the former colonies and on the frontier where President Jefferson’s policy of trading with them on credit to drive them into debt underlay ostensibly legal treaties, wherein selling their land settled the demand for debt payment. The millennia-old Indo-European cultural pattern of competition and its doppelgänger, conflict, had crossed the Atlantic and flourished.
The warrior spirit has been an important component of Afrikaner culture since its inception. The Boers’ heroic battle against the British during the Anglo-Boer War (1899-1902) for instance became an important part of the Afrikaner’s identity (Stapleton, 2010:106). During the so-called Border War (1966-1989), when South African conscripts fought against Communist expansion in South West Africa (now Namibia) and Angola, soldiering, masculinity and Afrikaner pride became inextricably entwined (Cock, 1989). After 1994, Afrikaners as a group have not been involved in violent conflicts (although some joined private military companies), but the warrior spirit survives in the Afrikaner’s primary sport, rugby, which is entwined with perceptions of masculinity, as discussed in Grundlingh (1994).
The Yamnaya culture relied on two innovative technologies in order to become a mobile and nomadic culture: the domesticated horse and the wheeled wagon. The earliest indication that horses became symbolically associated with humans dates to the early Yamnaya culture, around 4 500 BC, where they are found buried with humans (Anthony, 2007:201, 2023:20). These horses were the ancestors of modern horses, with the latter first appearing in the Don-Volga steppes around 2 200 BC (Anthony, 2023:20). After the Yamnaya expansion into Europe, horses became part of European culture. Indo-Europeans considered the horse to be sacred (Gimbutas, 1999:183), and numerous Viking Age burial sites include buried horses (Price, 2013:379-498). The Afrikaner’s association with horses can be seen in the numerous statues of Boer soldiers around South Africa, the revival of the horse by the Afrikaner Resistance Movement (Afrikaner Weerstandsbeweging or AWB), and in contemporary Afrikaner performances such as the Majuba Day commemoration in Orania or the Nooitgedachter horses on Orania’s currency, the Ora.
Wheeled vehicles were invented in the middle of the fourth millennium BC and the technology spread through the Pontic-Caspian steppe from 3 400 BC onwards (Anthony, 2023:21). Drawn by oxen, wheeled wagons made it possible for the Yamnaya to adopt a nomadic lifestyle, and Anthony (2023:21) calls the introduction of wheeled wagons an event “that partly defines the Yamnaya culture.” As a result of the ability to move possessions, the Yamnaya developed a society characterised by feasting and fighting (Anthony, 2023:21). Like horses, wagons also acquired symbolic significance, with numerous Yamnaya burial sites including wagons as part of grave goods (Anthony, 2023:28). Similarly to horses, wagons are also commonly found in Viking Age burials (Price, 2013:379-498), which indicates a symbolic significance. And as is the case with horses, the wagon is such a symbolic feature of the Afrikaner’s heritage that paintings and sculptures often depict wagons, and there is even the opportunity to overnight in a wagon in Orania.
It is notable that dogs were not eaten in the cultures of the Russian steppes (as opposed to some Asian cultures), and the ritual sacrifice of dogs at the site of Krasnosamarskoe in Russia may have been an initiation for young men to learn to kill humans (Anthony and Brown, 2017:139). As such, the bond between the Yamnaya and dogs was so strong that it facilitated using dogs as substitutes for humans. In the Germanic belief, wolves have special significance, e.g. as Óðinn’s pets (Freki and Geri), and dogs were regularly included in Viking Age burials (Price, 2013:379-498). In Afrikaner culture, dogs are prominent companions, with even a local breed, the boerboel (which is also the name of a popular clothing line that specifically markets to the Afrikaner).
Besides the warrior spirit, Kehoe (2012:12) mentions Indo-Europeans’ affinity for the number 3 (cf. three Norns, Fates and Moirae), which she argues survives linguistically in expressions such as “every Tom, Dick, and Harry”; “bell, book, and candle”; and “man, woman, and child”. In the Germanic belief, 9 (32) is “the magic number of the north, related to fertility, magic and royal cult” (Chaney, 1960:216). Óðinn hanged himself for nine days (Price, 2013:290), Freyr was worshipped at Uppsala every nine years by nine-day-long sacrificial rites (Gimbutas, 1999:196), and the Vølva had a council of nine wise women in pre-Christian Scandinavia (Gimbutas, 1999:119). Chaney (1960:207) highlights the trinities in the Germanic belief, such as Óðinn, Þórr, and Týr or Óðinn, Víli and Vé. In the Christian faith, the proclivity for the number 3 became the Holy Trinity, according to Kehoe (2012:12) and Chaney (1960:207). Gimbutas (1999:38) also discusses the triangle in the Neolithic European world, while Winterbourne (2004:156-157) notes the importance of this number in other beliefs. Gimbutas (1999:192) believes that the Norns were inherited by the Steppe newcomers from the Old Europeans, and if her analysis is correct, the importance of the number 3 may also date back further than the Yamnaya expansion. It is interesting to note that the number 3 also has special significance in Afrikaner culture, with the Driemanskap (Triumvirate) – consisting of Paul Kruger, Marthinus Wessel Pretorius and Piet Joubert – being prominent Afrikaner heroes (with a monument dedicated to them in Heidelberg), and the celebrated first Afrikaans poets were also known as the Driemanskap (Jan FE Celliers, Totius and C Louis Leipoldt). During apartheid, power was held through a combination of three institutions: The National Party, the Broederbond, and the Dutch Reformed Church.
Conclusion
Culture is not the product of a few decades or even centuries of development, but it rather develops over millennia. In the case of Northern European cultures, there is a measure of continuity from at least the Mesolithic to the present. The current study traced the Afrikaner’s genetic prehistory back to the Paleolithic, and showed how successive migrations of populations into Europe shaped Europeans’ physical attributes and culture. Special emphasis was placed on how vestiges of these migrations survived in European and in Afrikaner culture. Core issues associated with Afrikaner identity – language, whiteness, and a patriarchal social system – are directly inherited from the Yamnaya, and to a lesser extent from Neolithic farmers and WHG, while other cultural markers such as diet and the warrior spirit also show continuity from the Yamnaya to the present. In addition, important symbols of the Afrikaner – the wagon and the horse – also characterise the Yamnaya. Overall, although histories of the Afrikaner usually start in 1652, a surprising number of facets of the Afrikaner can be traced back much further.
Bibliography
Ahn, Y.-Y., Ahnert, S. E., Bagrow, J. P. and Barabási, A.-L. 2011. “Flavor network and the principles of food pairing,” Scientific Reports, 1:196. doi: 10.1038/srep00196.
Allentoft, M. E., Sikora, M., Refoyo-Martínez, A., Irving-Pease, E. K., Fischer, A., Barrie, W., Ingason, A., Stenderup, J., Sjögren, K.-G., Pearson, A., Sousa da Mota, B., Paulsson, B. S., Halgren, A. S., Schjellerup Jørkov, M. L., Demeter, F., Novosolov, M., Macleod, R., Sørensen, L., Nielsen, P. O., Henriksen, R. A. and Willerslev, E. 2022. “Population genomics of Stone Age Eurasia,” BioRxiv. doi: 10.1101/2022.05.04.490594.
Allentoft, M. E., Sikora, M., Sjögren, K.-G., Rasmussen, S., Rasmussen, M., Stenderup, J., Damgaard, P. B., Schroeder, H., Ahlström, T., Vinner, L., Malaspinas, A.-S., Margaryan, A., Higham, T., Chivall, D., Lynnerup, N., Harvig, L., Baron, J., Della Casa, P., Dąbrowski, P., Duffy, P. R. and Willerslev, E. 2015. “Population genomics of Bronze Age Eurasia,” Nature, 522(7555):167–172. doi: 10.1038/nature14507.
Anthony, D. W. and Brown, D. R. 2017. “The dogs of war: A Bronze Age initiation ritual in the Russian steppes,” Journal of anthropological archaeology, 48:134–148. doi: 10.1016/j.jaa.2017.07.004.
Anthony, D. W. 2007. The horse, the wheel, and language: how Bronze-Age riders from the Eurasian steppes shaped the modern world. Princeton: Princeton University Press.
Anthony, D. W. 2023. “The Yamnaya Culture and the invention of Nomadic pastoralism in the Eurasian steppes,” in Kristiansen, K., Kroonen, G., and Willerslev, E. (eds.) The Indo-European puzzle revisited: Integrating archaeology, genetics, and linguistics. Cambridge: Cambridge University Press:13–33.
Bousquette, J. and Salmons, J. 2017. “Germanic,” in Kapović, M. (ed.) The Indo-European languages. Oxon: Routledge:387–420.
Burger, J., Link, V., Blöcher, J., Schulz, A., Sell, C., Pochon, Z., Diekmann, Y., Žegarac, A., Hofmanová, Z., Winkelbach, L., Reyna-Blanco, C. S., Bieker, V., Orschiedt, J., Brinker, U., Scheu, A., Leuenberger, C., Bertino, T. S., Bollongino, R., Lidke, G., Stefanović, S. and Wegmann, D. 2020. “Low prevalence of lactase persistence in Bronze Age Europe indicates ongoing strong selection over the last 3,000 years,” Current Biology, 30(21):4307-4315.e13. doi: 10.1016/j.cub.2020.08.033.
Chaney, W. A. 1960. “Paganism to Christianity in Anglo-Saxon England,” Harvard Theological Review, 53(3):197–217. doi: 10.1017/S0017816000027012.
Chintalapati, M., Patterson, N. and Moorjani, P. 2022. “The spatiotemporal patterns of major human admixture events during the European Holocene,” eLife, 11. doi: 10.7554/eLife.77625.
Cock, J. 1989. War and Society: The Militarisation of South Africa. David Philip.
Cole, R. K. E. 2015. “Racial thinking in Old Norse literature: The case of the Blámaðr,” Saga Book, 39:21–40.
Cox, O. 1959. Caste, class and race. New York: Monthly Review Press.
Cox, S. L., Ruff, C. B., Maier, R. M. and Mathieson, I. 2019. “Genetic contributions to variation in human stature in prehistoric Europe,” Proceedings of the National Academy of Sciences of the United States of America. doi: 10.1073/pnas.1910606116.
Democratic Alliance 2021. DA files HRC complaint against Blade Nzimande for classifying Afrikaans as a “foreign” language. Available at: https://www.polity.org.za/article/da-files-hrc-complaint-against-blade-nzimande-for-classifying-afrikaans-as-a-foreign-language-2021-09-27 (Accessed: September 28, 2021).
Evershed, R. P., Payne, S., Sherratt, A. G., Copley, M. S., Coolidge, J., Urem-Kotsu, D., Kotsakis, K., Ozdoğan, M., Ozdoğan, A. E., Nieuwenhuyse, O., Akkermans, P. M. M. G., Bailey, D., Andeescu, R.-R., Campbell, S., Farid, S., Hodder, I., Yalman, N., Ozbaşaran, M., Biçakci, E., Garfinkel, Y. and Burton, M. M. 2008. “Earliest date for milk use in the Near East and southeastern Europe linked to cattle herding,” Nature, 455(7212):528–531. doi: 10.1038/nature07180.
Fu, Q., Posth, C., Hajdinjak, M., Petr, M., Mallick, S., Fernandes, D., Furtwängler, A., Haak, W., Meyer, M., Mittnik, A., Nickel, B., Peltzer, A., Rohland, N., Slon, V., Talamo, S., Lazaridis, I., Lipson, M., Mathieson, I., Schiffels, S., Skoglund, P. and Reich, D. 2016. “The genetic history of Ice Age Europe,” Nature, 534(7606):200–205. doi: 10.1038/nature17993.
Furtwängler, A., Rohrlach, A. B., Lamnidis, T. C., Papac, L., Neumann, G. U., Siebke, I., Reiter, E., Steuri, N., Hald, J., Denaire, A., Schnitzler, B., Wahl, J., Ramstein, M., Schuenemann, V. J., Stockhammer, P. W., Hafner, A., Lösch, S., Haak, W., Schiffels, S. and Krause, J. 2020. “Ancient genomes reveal social and genetic structure of Late Neolithic Switzerland,” Nature Communications, 11(1):1915. doi: 10.1038/s41467-020-15560-x.
Garnier, R., Sagart, L. and Sagot, B. 2017. “Milk and the Indo-Europeans,” in Robbeets, M. and Savelyev, A. (eds.) Language dispersal beyond farming. Amsterdam: John Benjamins:291–312.
Gimbutas, M. A. 1956. The prehistory of eastern Europe. Cambridge, Massachusetts: Peabody Museum.
Gimbutas, M. A. 1999. The Living Goddesses. Edited by M. R. Dexter. Berkeley: University of California Press.
Grasgruber, P., Cacek, J., Kalina, T. and Sebera, M. 2014. “The role of nutrition and genetics as key determinants of the positive height trend,” Economics and human biology, 15:81–100. doi: 10.1016/j.ehb.2014.07.002.
Gretzinger, J., Sayer, D., Justeau, P., Altena, E., Pala, M., Dulias, K., Edwards, C. J., Jodoin, S., Lacher, L., Sabin, S., Vågene, Å. J., Haak, W., Ebenesersdóttir, S. S., Moore, K. H. S., Radzeviciute, R., Schmidt, K., Brace, S., Bager, M. A., Patterson, N., Papac, L. and Schiffels, S. 2022. “The Anglo-Saxon migration and the formation of the early English gene pool,” Nature, 610(7930):112–119. doi: 10.1038/s41586-022-05247-2.
Grundlingh, A. 1994. “Playing for power? rugby, Afrikaner nationalism and masculinity in South Africa, c.1900–70,” The International journal of the history of sport, 11(3):408–430. doi: 10.1080/09523369408713871.
Haak, W., Lazaridis, I., Patterson, N., Rohland, N., Mallick, S., Llamas, B., Brandt, G., Nordenfelt, S., Harney, E., Stewardson, K., Fu, Q., Mittnik, A., Bánffy, E., Economou, C., Francken, M., Friederich, S., Pena, R. G., Hallgren, F., Khartanovich, V., Khokhlov, A. and Reich, D. 2015. “Massive migration from the steppe was a source for Indo-European languages in Europe,” Nature, 522(7555):207–211. doi: 10.1038/nature14317.
Hanel, A. and Carlberg, C. 2020. “Skin colour and vitamin D: An update,” Experimental Dermatology, 29(9):864–875. doi: 10.1111/exd.14142.
Hollfelder, N., Erasmus, J. C., Hammaren, R., Vicente, M., Jakobsson, M., Greeff, J. M. and Schlebusch, C. M. 2020. “Patterns of African and Asian admixture in the Afrikaner population of South Africa,” BMC Biology, 18(1):16. doi: 10.1186/s12915-020-0746-1.
Issa, L., Alghanim, F. and Obeid, N. 2018. “Analysis of food pairing in some eastern mediterranean countries,” in 2018 8th International Conference on Computer Science and Information Technology (CSIT), IEEE:167–172. doi: 10.1109/CSIT.2018.8486256.
Iversen, R. and Kroonen, G. 2017. “Talking Neolithic: Linguistic and archaeological perspectives on how Indo-European was implemented in Southern Scandinavia,” American Journal of Archaeology, 121(4):511–525. doi: 10.3764/aja.121.4.0511.
Jones, E. R., Gonzalez-Fortes, G., Connell, S., Siska, V., Eriksson, A., Martiniano, R., McLaughlin, R. L., Gallego Llorente, M., Cassidy, L. M., Gamba, C., Meshveliani, T., Bar-Yosef, O., Müller, W., Belfer-Cohen, A., Matskevich, Z., Jakeli, N., Higham, T. F. G., Currat, M., Lordkipanidze, D., Hofreiter, M. and Bradley, D. G. 2015. “Upper Palaeolithic genomes reveal deep roots of modern Eurasians,” Nature Communications, 6:8912. doi: 10.1038/ncomms9912.
Kassian, A. S., Zhivlov, M., Starostin, G., Trofimov, A. A., Kocharov, P. A., Kuritsyna, A. and Saenko, M. N. 2021. “Rapid radiation of the inner Indo-European languages: an advanced approach to Indo-European lexicostatistics,” Linguistics, 59(4):949–979. doi: 10.1515/ling-2020-0060.
Kehoe, A. 2012. Militant Christianity: An anthropological history. Palgrave Macmillan.
Kim, K.-J. and Chung, C.-H. 2016. “Tell me what you eat, and I will tell you where you come from: A data science approach for global recipe data on the web,” IEEE access : practical innovations, open solutions, 4:8199–8211. doi: 10.1109/ACCESS.2016.2600699.
Kristiansen, K., Allentoft, M. E., Frei, K. M., Iversen, R., Johannsen, N. N., Kroonen, G., Pospieszny, Ł., Price, T. D., Rasmussen, S., Sjögren, K.-G., Sikora, M. and Willerslev, E. 2017. “Re-theorising mobility and the formation of culture and language among the Corded Ware Culture in Europe,” Antiquity, 91(356):334–347. doi: 10.15184/aqy.2017.17.
Kristiansen, K. 2020. “The Archaeology of Proto-Indo-European and Proto-Anatolian: Locating the split,” in Serangeli, M. and Olander, T. (eds.) Dispersals and Diversification. Leiden: BRILL. doi: 10.1163/9789004416192_009.
Kroonen, G., Jakob, A., Palmér, A. I., Van Sluis, P. and Wigman, A. 2022. “Indo-European cereal terminology suggests a Northwest Pontic homeland for the core Indo-European languages,” Plos One, 17(10):e0275744. doi: 10.1371/journal.pone.0275744.
Kular, D. K., Menezes, R. and Ribeiro, E. 2011. “Using network analysis to understand the relation between cuisine and culture,” in 2011 IEEE Network Science Workshop, IEEE:38–45. doi: 10.1109/NSW.2011.6004656.
Lazaridis, I., Alpaslan-Roodenberg, S., Acar, A., Açıkkol, A., Agelarakis, A., Aghikyan, L., Akyüz, U., Andreeva, D., Andrijašević, G., Antonović, D., Armit, I., Atmaca, A., Avetisyan, P., Aytek, A. İ., Bacvarov, K., Badalyan, R., Bakardzhiev, S., Balen, J., Bejko, L., Bernardos, R. et al. 2022. “The genetic history of the Southern Arc: A bridge between West Asia and Europe,” Science, 377(6609):eabm4247. doi: 10.1126/science.abm4247.
Lazaridis, I. 2018. “The evolutionary history of human populations in Europe,” Current Opinion in Genetics & Development, 53:21–27. doi: 10.1016/j.gde.2018.06.007.
Mallory, J., Dybo, A. and Balanovsky, O. 2019. “The impact of genetics research on archaeology and linguistics in Eurasia,” Russian journal of genetics, 55(12):1472–1487. doi: 10.1134/S1022795419120081.
Mallory, J. P. 1989. In search of the Indo-Europeans: language, archaeology and myth. London: Thames & Hudson.
Margaryan, A., Lawson, D. J., Sikora, M., Racimo, F., Rasmussen, S., Moltke, I., Cassidy, L. M., Jørsboe, E., Ingason, A., Pedersen, M. W., Korneliussen, T., Wilhelmson, H., Buś, M. M., de Barros Damgaard, P., Martiniano, R., Renaud, G., Bhérer, C., Moreno-Mayar, J. V., Fotakis, A. K., Allen, M. and Willerslev, E. 2020. “Population genomics of the Viking world,” Nature, 585(7825):390–396. doi: 10.1038/s41586-020-2688-8.
Marnetto, D., Pankratov, V., Mondal, M., Montinaro, F., Pärna, K., Vallini, L., Molinaro, L., Saag, L., Loog, L., Montagnese, S., Costa, R., Estonian Biobank Research Team, Metspalu, M., Eriksson, A. and Pagani, L. 2022. “Ancestral genomic contributions to complex traits in contemporary Europeans,” Current Biology, 32(6):1412-1419.e3. doi: 10.1016/j.cub.2022.01.046.
Maylam, P. 2020. South Africa’s Racial Past: The History and Historiography of Racism, Segregation, and Apartheid. London: Routledge.
Mittnik, A., Wang, C.-C., Pfrengle, S., Daubaras, M., Zariņa, G., Hallgren, F., Allmäe, R., Khartanovich, V., Moiseyev, V., Furtwängler, A., Andrades Valtueña, A., Feldman, M., Economou, C., Oinonen, M., Vasks, A., Tõrv, M., Balanovsky, O., Reich, D., Jankauskas, R., Haak, W. and Krause, J. 2017. “The genetic history of northern europe,” BioRxiv. doi: 10.1101/113241.
Morez, A., Britton, K., Noble, G., Günther, T., Götherström, A., Rodríguez-Varela, R., Kashuba, N., Martiniano, R., Talamo, S., Evans, N. J., Irish, J. D., Donald, C. and Girdland-Flink, L. 2022. “Imputed genomes and haplotype-based analyses of the Picts of early medieval Scotland reveal fine-scale relatedness between Iron Age, early medieval and the modern people of the UK,” BioRxiv. doi: 10.1101/2022.08.01.502257.
Mpeta, B., Fourie, J. and Inwood, K. 2018. “Black living standards in South Africa before democracy: New evidence from height,” South African journal of science, 114(1/2):1–8. doi: 10.17159/sajs.2018/20170052.
Myburgh, J., Staub, K., Rühli, F. J., Smith, J. R. and Steyn, M. 2017. “Secular trends in stature of late 20th century white South Africans and two European populations,” Homo: internationale Zeitschrift fur die vergleichende Forschung am Menschen, 68(6):433–439. doi: 10.1016/j.jchb.2017.10.001.
Neparáczki, E., Maróti, Z., Kalmár, T., Kocsy, K., Maár, K., Bihari, P., Nagy, I., Fóthi, E., Pap, I., Kustár, Á., Pálfi, G., Raskó, I., Zink, A. and Török, T. 2018. “Mitogenomic data indicate admixture components of Central-Inner Asian and Srubnaya origin in the conquering Hungarians,” Plos One, 13(10):e0205920. doi: 10.1371/journal.pone.0205920.
Ngcobo, P. N., Nyati, L. H., Norris, S. A. and Pettifor, J. M. 2021. “The age at which adult height is achieved during adolescence in the Birth to Twenty Cohort, Johannesburg, South Africa,” South African Journal of Child Health, 15(2):60–65.
Olalde, I., Brace, S., Allentoft, M. E., Armit, I., Kristiansen, K., Booth, T., Rohland, N., Mallick, S., Szécsényi-Nagy, A., Mittnik, A., Altena, E., Lipson, M., Lazaridis, I., Harper, T. K., Patterson, N., Broomandkhoshbacht, N., Diekmann, Y., Faltyskova, Z., Fernandes, D., Ferry, M. and Reich, D. 2018. “The Beaker phenomenon and the genomic transformation of northwest Europe,” Nature, 555(7695):190–196. doi: 10.1038/nature25738.
Olander, T. 2019. “The Indo-European homeland: Introducing the problem,” in Olsen, B. A., Olander, T., and Kristiansen, K. (eds.) Tracing the Indo-Europeans. New evidence from archaeology and historical linguistics. Oxford: Oxbow Books:7–34.
Olander, T. (ed.) 2022. The Indo-European Language Family. A Phylogenetic Perspective. Cambridge: Cambridge University Press.
Pickersgill, B. 2004. “Spices,” in Prance, G. T. and Nesbitt, M. (eds.) The Cultural History of Plants. New York: Routledge:153–172.
Posth, C., Yu, H., Ghalichi, A., Rougier, H., Crevecoeur, I., Huang, Y., Ringbauer, H., Rohrlach, A. B., Nägele, K., Villalba-Mouco, V., Radzeviciute, R., Ferraz, T., Stoessel, A., Tukhbatova, R., Drucker, D. G., Lari, M., Modi, A., Vai, S., Saupe, T., Scheib, C. L. and Krause, J. 2023. “Palaeogenomics of Upper Palaeolithic to Neolithic European hunter-gatherers,” Nature, 615(7950):117–126. doi: 10.1038/s41586-023-05726-0.
Price, N. 2013. The Viking Way: Magic and mind in late Iron Age Scandinavia. Oxford: Oxbow Books.
Racimo, F., Woodbridge, J., Fyfe, R. M., Sikora, M., Sjögren, K.-G., Kristiansen, K. and Vander Linden, M. 2020. “The spatiotemporal spread of human migrations during the European Holocene,” Proceedings of the National Academy of Sciences of the United States of America, 117(16):8989–9000. doi: 10.1073/pnas.1920051117.
Renfrew, C. 1989. Archaeology and language. The Puzzle of Indo-European Origins. London: Penguin Books.
Rosenstock, E., Ebert, J., Martin, R., Hicketier, A., Walter, P. and Groß, M. 2019. “Human stature in the Near East and Europe ca. 10,000–1000 BC: its spatiotemporal development in a Bayesian errors-in-variables model,” Archaeological and anthropological sciences, 11(10):5657–5690. doi: 10.1007/s12520-019-00850-3.
Sajadmanesh, S., Jafarzadeh, S., Ossia, S. A., Rabiee, H. R., Haddadi, H., Mejova, Y., Musolesi, M., Cristofaro, E. D. and Stringhini, G. 2017. “Kissing cuisines: exploring worldwide culinary habits on the web,” in Proceedings of the 26th International Conference on World Wide Web Companion – WWW ’17 Companion. New York, New York, USA: ACM Press:1013–1021. doi: 10.1145/3041021.3055137.
Segurel, L., Guarino-Vignon, P., Marchi, N., Lafosse, S., Laurent, R., Bon, C., Fabre, A., Hegay, T. and Heyer, E. 2020. “Why and when was lactase persistence selected for? Insights from Central Asian herders and ancient DNA,” PLoS Biology, 18(6):e3000742. doi: 10.1371/journal.pbio.3000742.
Senekal, B. A. 2019. “Ras en Afrikaneretnisiteit: ʼn Kwantitatiewe ondersoek na huidige opvattinge,” Ensovoort, 40(8).
Senekal, B. A. 2022. “Wat eet Afrikaners? ’n Verkenning van die bestanddeel-resep-netwerk in Afrikaanse resepte,” LitNet Akademies (Natuurwetenskappe), 19(3):648–670.
Senekal, B. A. 2023. “’n Visuele verkenning van Suid-Afrika se posisie in die globale bestanddeel-resep-netwerk,” forthcoming.
Serrano, J. G., Ordóñez, A. C. and Fregel, R. 2021. “Paleogenomics of the prehistory of Europe: human migrations, domestication and disease,” Annals of Human Biology, 48(3):179–190. doi: 10.1080/03014460.2021.1942205.
Shishlina, N., Sevastyanov, V. and Hedges, R. E. M. 2012. Isotope ratio study of Bronze Age samples from the Eurasian Caspian Steppes. Berlin: De Gruyter.
Skoglund, P. and Mathieson, I. 2018. “Ancient genomics of modern humans: the first decade,” Annual Review of Genomics and Human Genetics, 19:381–404. doi: 10.1146/annurev-genom-083117-021749.
Stapleton, T. J. 2010. A military history of South Africa: From the Dutch-Khoi Wars to the end of apartheid. Santa Barbara: Praeger.
Stolarek, I., Handschuh, L., Juras, A., Nowaczewska, W., Kóčka-Krenz, H., Michalowski, A., Piontek, J., Kozlowski, P. and Figlerowicz, M. 2019. “Goth migration induced changes in the matrilineal genetic structure of the central-east European population,” Scientific Reports, 9(1):6737. doi: 10.1038/s41598-019-43183-w.
Wilkin, S., Ventresca Miller, A., Fernandes, R., Spengler, R., Taylor, W. T.-T., Brown, D. R., Reich, D., Kennett, D. J., Culleton, B. J., Kunz, L., Fortes, C., Kitova, A., Kuznetsov, P., Epimakhov, A., Zaibert, V. F., Outram, A. K., Kitov, E., Khokhlov, A., Anthony, D. and Boivin, N. 2021. “Dairying enabled Early Bronze Age Yamnaya steppe expansions,” Nature, 598(7882):629–633. doi: 10.1038/s41586-021-03798-4.
Willemse, H. 2018. “The hidden histories of Afrikaans,” in Netshitenzhe, J. (ed.) Whiteness, Afrikaans, Afrikaners: Addressing post-apartheid legacies, privileges and burdens. Woodmead, Johannesburg: Mapungubwe Institute for Strategic Reflection (MISTRA):115–130.
Winterbourne, A. 2004. When the Norns have spoken: Time and fate in Germanic Paganism. Madison, N.J: Fairleigh Dickinson University Press.
Zhu, Y.-X., Huang, J., Zhang, Z.-K., Zhang, Q.-M., Zhou, T. and Ahn, Y.-Y. 2013. “Geography and similarity of regional cuisines in China,” Plos One, 8(11):e79161. doi: 10.1371/journal.pone.0079161.